Thursday, February 4, 2010
Did the Moa's ancestor fly to New Zealand?


The group of birds to which the moa belonged, the ratites, have long fascinated evolutionary biologists. All the ratites are flightless (though, as we'll see they are related to the quail-like tinamous which can fly passably well) and all the major landmasses that had their start in Gondwana had at least one ratite species before humans arrived on the scene. Africa has ostriches, South America the rhea, Australians have emus and cassowary, Madagascar had the Elepahant bird and New Zealand lost the moa but retains the kiwi. The far flung distribution of the ratites and their apparent lack of ability to disperse between continents has led to them being put forward as a classic example of an idea called vicariance biogeography in which the evolutionary history of a group is driven by the geological history of the land on which they live.
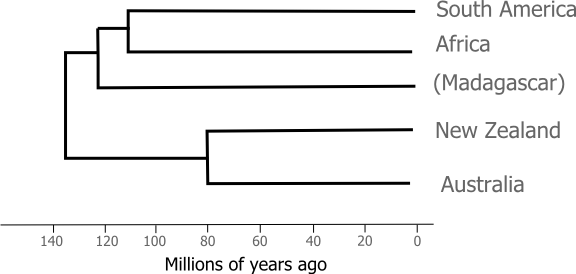
For vicariance biogeographers the evolution of the ratites was driven by the movement of the continents. The ancestor of all modern ratites was a flightless bird living in Gondwana and as each new continent split and rifted away from the super continent it took with it a population of ratites which adapted to the ecological changes brought on by their continent's journey: cassowaries in the Wet Tropics of Australia, ostriches on the African Savannah, rhea on the Pampas. It's certainly a nice story, but science has a way of ruining nice stories. The role of vicariance of evolution in the ratites was put to the test once we became able to use molecular evidence to reconstruct the relationships between species. If the geologically driven sketch of ratite evolution I presented above is right then the pattern of branching we find among ratites from different continents should match the order in which we know the continents broke up, something like this:
In 2001 Alan Cooper and colleagues sequenced the entire mitochondrial genome (some 12 000 bases of DNA) from representatives of each of the extant ratites and, remarkably, two species of moa. The long, careful process of retrieving DNA sequences from sub-fossil bones deserves a post of its own but for the sake of this article we only need to know what Cooper et al found when they used that DNA to recover the the relationships between ratite species.

The species in bold text above don't fit the pattern that we'd expect from geology alone. If ratite relationships simply reflected the Gondwanan breakup we'd expect to see ostriches grouping with rheas (and apart from the other ratites). New Zealand's two ratite orders are even more surprising, the kiwi lineage is more closely related to the Australian ratites than it is to the moa species. When combined with a molecular clock analysis Cooper et al. concluded that modern kiwis are the descendants of ancient immigrants hailing from either Australia or islands in the Lord Howe Rise (which have since submerged). In order to explain that trans-tasman dispersal the authors reached for the last resort of the desperate biogeographer and invoked a land bridge for which there is little geological evidence. In fact, as we'll see it now seems more likely that the ancestors of the kiwi and the moa flew to New Zealand.
Even with the mitochondrial phylogeny of the group published there was considerable room for uncertainty in how the ratites related to each other. The underlying shape of ratite tree makes it particularly difficult to accurately recover with phylogenetic methods. When we use DNA sequences to estimate a phylogenetic tree we need to find species that share mutations that have accrued during the evolution of the group we're looking at. The branches that relate the different ratite species are relatively short, so there was little time for mutations that set related groups apart from more distantly related ones to accrue. Even worse, the branches that reach to the modern species (the tips of the tree) are very long meaning there has been a lot of time to any mutations that did accrue in those critical short branches to be overwritten*. There are three approaches to dealing with this problem - sequence more genes (since each unlinked gene acts as a separate witness to the evolution of the group), sequence more samples (especially if doing so breaks up a long branch) or use a better model for the way mutations accrue in the genes you are studying. People have tried all three methods to get a better look at ratite evolution. Last year a group centred around the Field Museum in Chicago published a mutli-gene phylogeny of all birds that contained a big surprise for ratite evolution- the most recent common ancestor of all ratites flew.

As long as the ratites grouped together in a phylogeny it was reasonable to assume that they all inherited their flightlessness from the common ancestor of the group. The Field Museum study found that, in fact, the flying tinomous fit right in the middle of the flightless ratites. So, either the most recent common ancestor of the ratites and the tinamous flew and ratite lineages have subsequently lost that ability at least three times or that ancestor was grounded and the tinamous have rediscovered flight. In vertebrates the evolution of true flight has happened three times (in bats, pterosaurs and birds) while there are hundreds of examples of birds that have given up on flying. Moreover, a group of flying birds that are prone to flightlessness is hardly anything new - at least 30 species of rail (including our own weka and takahe) have taken to life on the ground. Given the ways the odds are stacked towards losing flight it seems probable the common ancestor that relates tinamous and ratites flew. The Field Museum study didn't include any moa species and didn't attempt any molecular dating so it's hard to see just how the ancestors of the kiwi and the moa made it to New Zealand
A new study (I knew I'd get to it eventually) published in Systematic Biology throws some light on the New Zealand ratite story. Matt Philips and a team of researchers from the Alan Wilson Centre at Massey University took another look at the mitochondrial dataset used in Cooper et al's 2001 study by adding more kiwi species and using models of DNA evolution that avoid some of the pitfalls of the ratite phylogeny's difficult shape. The new ratite tree and a molecular clock analysis based on that tree confirm the idea of multiple loses of flight in the ratites and add a new finding - the closest living relatives of our giant moa are the quail-like tinamous:
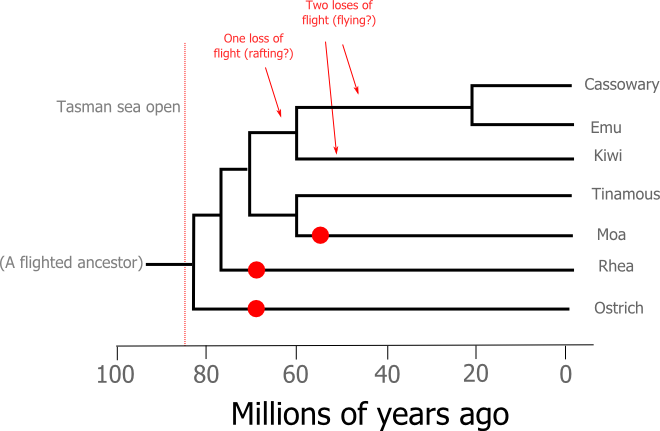
So what does the new understanding of ratite relationships mean for our ideas about the origins of New Zealand's ratites? The molecular clock doesn't quite tick with the regularity of a stopwatch, so there is a good deal more uncertainty in the timing of the events presented above than the precisely defined nodes suggest. Still, even with the uncertainty of molecular dating taken on board we can safely say that both the New Zealand ratite lineages departed from their closest relatives after the Tasman Sea opened up.
The revelation that the tinamous are the moa's closest living relatives suggests that the moa had ancestors that could fly. So, it seems the first proto-moa to arrive in New Zealand flew, or more likely was blown, here from Antarctica. Antarctica? It still seems amazing to me but 30 million years ago Antartica was still attached to South America and, without the circumpolar current to isolate from the world, was a relatively verdant continent. We know from fossils that Antarctica supported southern beech forests (still found in Chile, New Zealand and Australia) and marsupial mammals (strangely absent in New Zealand but still present in Australia and South America) so it's no great stretch to propose the representatives of another Gondwanan group lived there. Antarctica certainly seems like a more likely jumping off point for dispersing proto-moa than South America, but either way it certainly seems they made it here under their own power.
The mode of dispersal for the kiwi's ancestor is a little less clear. As we've seen we can be quite sure that they arrived in New Zealand after the Tasman Sea opened up and there is really no good evidence that there was ever a land-bridge across that sea. We can probably rule out walking. If we disregard the problem of dispersal for a second the simplest way to explain the distribution of flightlessness on the Phillips et al phylogeny is with a single loss of flight in an ancestor shared by kiwis and the Australian ratites. Under that scenario the kiwi would, presumably, have had to raft to New Zealand. Alternatively, given that we've seen the ratites seem to have an inbuilt propensity to becoming flightless we might imagine that the common ancestor shared by the kiwi and the Australian ratites could fly and each lineage has since lost that ability. In this case the kiwi could simply have flown from Australia to New Zealand (a journey that storms frequently inflict on Australian birds today). Without sufficiently old ratite fossils from either country it's hard to choose one scenario over the other.Long range dispersal by rafting is probably an important force in biogeography but if I was forced to make a bet I'd put my money on ancient flying kiwis.
The radical rethink of ratite evolution that a decade of molecular phylogenetics has forced on us raises a lot of interesting questions. What it is it about the ratite body plan, development or behaviour that makes them so prone to flightlessness? Is that repeated loss of flight, and consequent lack of pressure to keep their weight down, enough to explain the trend towards gigantism? The authors of the most recent paper suggest both trends might be explained by ratites on each continent filling the ecological niches left by the extinction of the dinosaurs. The dates on their tree are certainly consistent with the idea that each ratite lineage independently took to the ground 65 million years ago but without more fossils and more precise dates for each split it's very hard to test the idea further. I'm sure the story of ratite evolution has more surprises for us to uncover.
Links to the primary literature are provided below but you should also check out Simon Collins excellent piece in the Herald and Mike Dickison, who got his PhD studying giant flightless birds and wrote about the idea that ratites flew to New Zealand way back in 2007.
Cooper A, Lalueza-Fox C, Anderson S, Rambaut A, Austin J, & Ward R (2001). Complete mitochondrial genome sequences of two extinct moas clarify ratite evolution. Nature, 409 (6821), 704-7 PMID: 11217857 Harshman, J., Braun, E., Braun, M., Huddleston, C., Bowie, R., Chojnowski, J., Hackett, S., Han, K., Kimball, R., Marks, B., Miglia, K., Moore, W., Reddy, S., Sheldon, F., Steadman, D., Steppan, S., Witt, C., & Yuri, T. (2008). Phylogenomic evidence for multiple losses of flight in ratite birds Proceedings of the National Academy of Sciences, 105 (36), 13462-13467 DOI: 10.1073/pnas.0803242105 Phillips, M., Gibb, G., Crimp, E., & Penny, D. (2009). Tinamous and Moa Flock Together: Mitochondrial Genome Sequence Analysis Reveals Independent Losses of Flight among Ratites Systematic Biology, 59 (1), 90-107 DOI: 10.1093/sysbio/syp079Labels: biogeography, evolution, might interest someone, moa, new zealand, phylogenetics, ratites, sci-blogs, science
11 Comments:


jerseys cheap

All content not otherwise marked is licensed under a Creative Commons Attribution-ShareAlike 3.0 Unported License.