
Sunday, December 19, 2010
Sunday Spinelessness - Updates
I'm going to round out Sunday Spinelessness for 2010 with a few updates from the creatures that have featured here over the year. But, since this is going to be last post here till the first week of January or so, I'm going to start by taking just a moment to wish everyone who has commented on, linked to, tweeted about or just read The Atavism this year a merry Christmas and an enjoyable break over the new year. I enjoy writing these posts for their own sake, but it's much more satisfying to be part of a community (no matter how small a part!) of people that are interested in the same ideas I am. Thanks.
Right, now own with the spinelessness. The little green spiders I've been following, and sacrificing in the name of science are still going strong. Since the last time I wrote about them I've seen them grab prey for the first time (confirming the suspicion that they are ambush predators) and seen males fight for the right to mate with a female. Sadly, I didn't photograph either of these events (well, not in a way that I'd want to show other people, but there are descriptions of each tucked away in my notebook). I can, however, show you the result of those matings. About three weeks after the first one I started spotting these across the agapanthus:

At last count there were about five separate leaves sporting one of these egg sacs and a watchful mother. One very dedicated mum-to-be actually has three on the go at the moment!
I'll have to wait a while to see the next generation of the little green spiders, but the leaf-veined slugs that featured along side them here have already moved into the next stage

I haven't seen a full-sized slug in a long time. But these immature ones are out and about every evening, I'll have a little more to say about them in the new year.
The pittasporum "pysllids" are still dancing, but no longer at plague proportions. There's no respite for the pittasporum though, now it's being devoured by leaf roller moths. If I cared more about the tree I might spray it with insecticide but while its pests are feeding animals as wonderful as this spider (anyone have a clue on an ID?) I'm happy to let them have it:

Finally, the bumblebee nest at the bottom of the garden appears to be enjoying the run of hot weather we've had here. Some of the bees in our backyard (no necessarily the ones that live at the bottom, since bumble bees can forage several kilometres from their base) have taken up a slightly odd habit. On a couple of warm evenings I've popped outside and found bumblee bees asleep inside foxglove flowers:

No doubt the little tubes make a nice warm place for a worker to spend the night if they are caught out too late to make it home. Apparently males, which are only produced towards the end of summer, will often do the same trick once they've left the nest and are spending their daytime waiting for a virgin queen to pass by. I think we really have to end with a bumble bee bum sticking out of a flower:

Labels: arachnophilia, bee, blog blogging, bumblebee, environment and ecology, leaf veined slugs, sci-blogs, spider, sunday spinelessness
Tuesday, December 14, 2010
Not so flash short fiction
As I had predicted, I didn't make the short list for New Scientist's flash fiction competition. Here's what I had entered, reproductive health a Lamarkian world:
Tuesday.
He closed the door, put the pottle down and sighed. Tuesdays were the worst. He’d adjusted to the early morning runs the late night saunas, he’d even come to like the all-curry diet but he couldn’t get used to this.
Before, when this had started, he’d boil from one emotion to the next on Tuesdays. He'd be angry at Janine for wanting to do it this way; angry at his parents for leaving him with this; disgusted by this harshly lit white room, its sterile lino floor and the list of men who’d used it before him. Today he didn’t even care as his flipped through the same set of magazines again. What was the point? Even if he could change Janine’s mind he wouldn’t, and his parents couldn’t have known their choices where effecting anyone by themselves. Hell, he’d got to thirty without finding out, on the first pre-conception check up:
CFTR: 46% (+/- 3%) germules abnormal
His parents hadn't exercised, the doctor explained, so the germules that would have helped them breathe and sweat efficiently didn't get used. As a result, he'd inherited a suite of derelict germules , the most dangerous of which was one called CFTR. He had enough healthy CFTR to get by, but because Janine had the normal background level of broken ones (about 4%) there was a chance their children would have cystic fibrosis. Once the counsellor explained the disease it was obvious they should avoid it. They might have created and screened embryos, only implanting healthy ones, but Janine couldn't destroy the ones that didn't make the grade.
So, his life had become almost entirely about his germules: chilli, running and saunas to exercise them and Tuesdays to test them. That meant a bleak ten minutes in this bright room, after which he handed Louise, the unnerving cheery nurse, his sample. He couldn’t even muster embarrassment at this transaction anymore, so he avoided the waiting room and stuck around for the verdict:
"It's definitely working, 23%, that's half way to the target! See you next Tuesday!"
Sunday, December 12, 2010
Sunday Spinelessness - What makes a bee get busy
I've spend quite a lot of time peering into this little pile of weeds recently.

That's not because I'm particularly proud of our ability to grow some of the most extensive chickweed and onion weed beds in town. Those weeds are covering the entrance to a bumblebee nest.

I don't know what it is about them, but watching big, fat buzzy bumblebees fly about never fails to make me happy. So every now and again I wander down to check in on the nest and watch them going about their business. Which got me to thinking about a pretty cool paper I read a while ago. When Ralph Stelzer took bumblebee nests from The Netherlands to Northern Finland, where it's light for weeks on end during the summer months, the bees kept to their normal cycle of foraging during the "daytime" hours and sleeping under the midnight sun. So bumblebees obviously have some sort internal of body clock that isn't reset by the length of the day. But, I wondered, what else contributes to the rate at which the workers go out and forage? There is actually quite a literature just that topic, but a couple of weeks in the field or the lab can often save minutes in the library so I decided to try and find out for myself.
For the last month, whenever I've been working from home I've popped down to the nest every two hours or so (that's about when I need a break from thinking anyway) and counted how many workers fly in or out of the nest in a three minute period. I'm going to use that measure of nest activity as my "response variable", the data that I am trying to find an explanation for, in a statistical analysis. What "predictor variables" should I use to try an explain nest activity? We know from Stelzer's paper that bumblebees have a body clock, so time of day is an obvious predictor to include. Similarly, I know that bumblebees can only fly when their wing muscles are around 30°C and the nectar they forage is going to be more abundant when the sun is shining and it's warm, so ambient temperature and solar energy might predict when a bumblebee can fly. I don't have weather station in the backyard, but there is one about a kilomertre away so I used that to estimate both the temperature and the solar irradiance (the amount energy from the sun reaching the ground) at home. I'm pretty sure the nest was new when I first started observing it, so it might still have been building up its workforce over the first few days, in which case the day on which a record was made might contribute to the number of flights recorded. Finally, I actually got out and counted bees on one rainy day so I should include "rainy or not" as a predictor.
Now comes the most important step in any analysis, taking a look at the data. How do all those variables relate to the number of bees making their way to and from the nest?
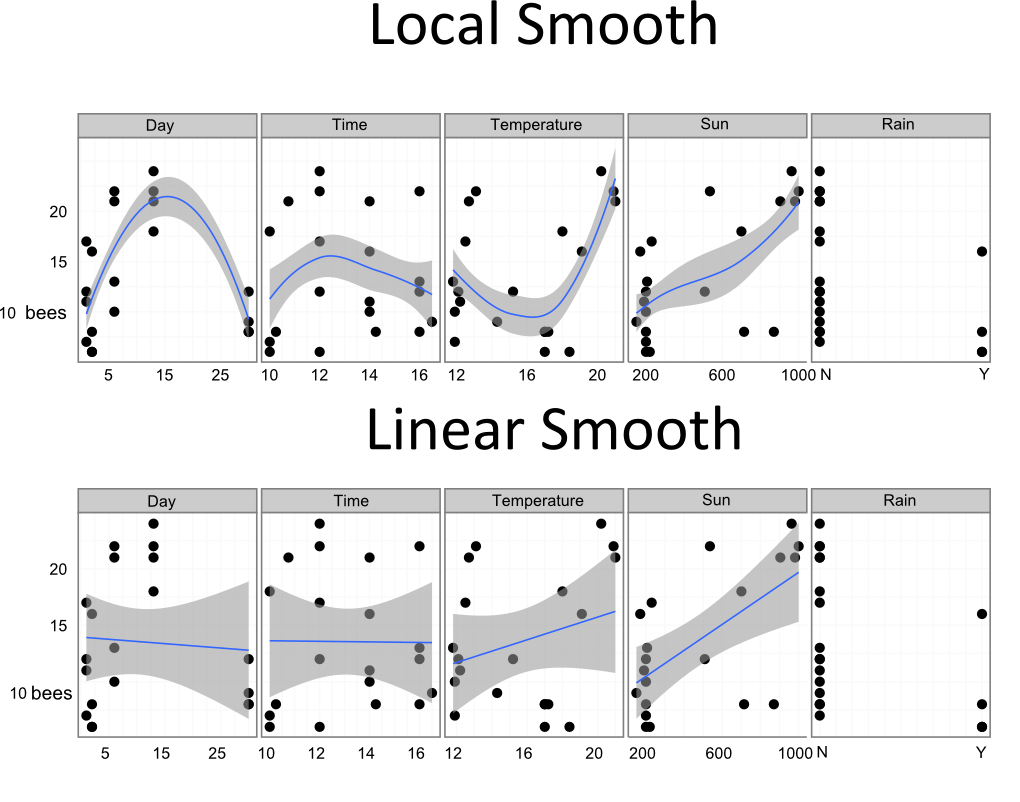
You can see a few relationships in the data, and some definite problems in the way I collected it. For one there is the massive gap between the second to last day and the final one. If I had made my records at more regular intervals I would have a better idea if that last day was an outlier if the local smooth's trajectory, with a peak in the middle, is the real pattern. Nevertheless, there are some clear patterns. Both temperature and solar irradiance seem to get bumblebees out there collecting nectar, and activity might peak around the middle of the day.
Having eye-balled the data, it's time to build a model to explain. I'm going to use a method called a generalized linear model (GLM). I realise it's a Sunday, and no one wants a stats leason on their day off, so here's the super-condensed summary of what the GLMs can do. They are really just an extension of the "plot some points and put a straight line through them" regressions displayed above. You include all the variables you think might contribute to the data you are trying to explain, then you can use some nifty tools to select a model that includes only those variables that are important predictors. In this case, we also need to explain that the "day", "time" and "temperature" variables might be better modeled as a curve rather than a straight line. You can probably remember from high school maths that the way to turn a straight line where y = x into a curved parabola is to square x, and to include a curved response for "day" we just include "day squared" in our GLM. So, our slightly terrifying looking starting point is a model like this:
number of bees = day + day2 + time + time2 + temp + temp2 + sun + rain
When we apply the nifty tools I talked about above to get rid of useless predictors, we get a less complex model that retains "rain", "temperature", "sun" and "day2" but gets rid of the rest.
log(number of bees)* = 1.83 + 0.05 . temp + 0.001 . sun + 0.001 . day2 - 0.6 . rain
The size of the numbers in those equations doesn't tell us much about their relative importance, since all the predictor variables are in different units. Instead, we need to look at how much of the variation in bee activity each of the retained variables explain:
| Variable | Percent variation explained** |
| day2 | 27 |
| rain | 14 |
| sun | 10 |
| temp | 10 |
So it seems the curved relationship between the day of observation and the number of bees explains more of the variation in the data that I collected than anything else. That's kind of annoying, since I'm not sure if the curved data with its peak of activity really descibes what's going on at the nest. Other than that long-term effect, we can see that bees fly when it's sunny dry and warm. Clearly I'm preparing this for Nature right now.
Of course, statistical models can only tell us how to explain the data they are given to test. My data is actually pretty crumby, estimating over-all activity using 3 minutes of data fits my schedule but it makes the estimate very rough. I only recorded between 10am and 4pm, even though bumblebees fly from dawn till dusk. I've also since learned that humidity is an important driver for foraging behavior and that variable isn't included in the model above. If you were looking for lesson in this post (I wasn't trying to write one) then it would be that any statistical analysis is only as good as the data you collect. If I was really serious about uncovering what made bumblebees tick, and I have enough time to do the question justice, then I'd been spending a lot more than 3 minutes on each record, and I'd be making a lot more then four records a day.
* The number of bees has to be log-transformed because, being counted data, it doesn't follow a nice bell-curve which is a requirement for a lot of statistical tests.
** Yes, my fellow stats geeks. This is a poisson-family GLM so it's actually a table of deviance but I thought this title better explained the what the numbers meant.
The Finnish bumblebee study is
Stelzer, R. & Chittka, L., 2010. Bumblebee foraging rhythms under the midnight sun measured with radiofrequency identification. BMC Biology, 8(1), 93.
Labels: bumblebee, geekery, sci-blogs, statistics, sunday spinelessness
Sunday, December 5, 2010
Sunday Spinelessness - The origin and extinction of species
I don't use these pages to write about my own work very much, partly because it's not yet published and partly because I write about that all day as it is. The shortest answer I can provide to the question "what do you do" is "I use genetic tools to study evolution" and I guess that makes me an evolutionary geneticist. You can split the people that work in our field into two groups: there are biologists that are really interested in a group of organisms and have learned some genetics to help their study of them, and there are people who are interested in a particular question and have chosen their study organisms to suit. I'm very much of the second sort, and like most people in that group I've caught myself saying "I'm interested in the questions, not the animals". Paraphrased, that becomes something like, "Oh sure, I study Pacific land snails, but for all I care they're just little bags of genes that help me answer questions". But that's a lie. You can't work on animals without having them effect you. When I started my PhD I had no particular love of snails, but now I'm a complete snail fan-boy and I frequently find myself preaching on the wonders of life as a terrestrial mollusc to people whose only mistake was to ask me what I do for a living. Did you know most slugs retain the remnants of their shells? Or that almost all snail shells coil to the right? Or that mating in many land snail species only proceeds after one snail has stabbed the other with a "love dart"? A couple of weeks ago I was recounting the the sad tale of The Society Islands partulids to someone I'd met three minutes earlier, and today I'm going to tell you that story (though, of course you have an advantage over the first recipient of the story, since you don't have to read this crap)
Believe it or not, land snails are one of the characteristic animals of Pacific Islands. Anak krakatau is so young it's still smoldering, and it has a native land snail species and Rapa nui (Easter Island), which is arguably the most isolated island in the Pacfic, had its own land snail fauna back when it had forests. It's not entirely clear how these unlikely colonists get to islands. Darwin was so interested in the question* he, ever the experimentalist, stuck snails to ducks' feet to see if they'd survive an inter-island journey. Birds have been shown to carry snails great distances, but wind blown leaves are probably a more common mode of conveyance. We might not know exactly how snails get to islands, but we know what happens once they establish themselves. The land snails of the Pacific include some of the most outrageous explosions of diversity in the biological world. Chief among these evolutionary radiations were the partulid snails of The Society Islands (the French Polynesian archipelago that includes Tahiti). Partulids are very elegant tree snails that form part of the land snail fauna across most of Polynesia, in the Societies they made up most of the land snail fauna. In total, the tiny islands had 58 species of these snails with each of the main islands have their own endemic forms.

A plate from Crampton's monograph on the partulids of Moorea
The Society Islands' land snails were a marvel all by themselves, but they were also an extraordinary resource for scientists. The first person to seriously take up their study was the American embryologist and evolutionary biologist Henry Crampton. Crampton was working at the turn of the 20th century, a time in which the mechanisms underlying genetics and evolution were very much up for debate, and he hoped Tahitian and Moorean partulids could help set the story straight. Crampton's monogrpahs are famous (at least among people that spend thier lives thinking about snails) for their detail. He collected and measured over two hundred thousand shells, then calculated summary statistics for each species, each site and each measurement. By hand. To eight decimal places.

Those massive tables (there are more than 100 pages of them in the Moorean monograph) might seem like an old-fashioned, descriptive, way to do biology. But in many ways Crampton was ahead of his time. For one, he was a Darwinist when not every evolutionist was. By the end of the 19th century Darwin had convinced the world of the fact evolution had happened, but relatively few naturalist bought his theory of how evolutionary change happened. The anti-Darwinian theories that prospered during the so called "eclipse of Darwinism" placed very little importance on the variation within species. The orthogenesists and the lamarckians thought evolution had a driving force, pushing species towards perfection. In their scheme variation within a species was deviance from the mainstream of evolution and was quckly stamped out by natural selection (which they didn't deny, they just said it couldn't be a creative force). Similarly, saltationists thought large-scale evolutionary changes occurred in a single generation, and the small changes you see in populations were of no consequence in the grand scheme of evolution. Crampton realised that, in a Darwinian world, variation within populations was the raw material of evolution. He was obsessive about measuring his shells because he knew could use the data he was recording to understand where species came from. In particular, we was able to show that isolated populations of the same species varied from each other. That finding that makes sense in light of Darwin's theory, since species arise from populations evolving away from each other; but is harder to fit into progressive theories of evolution, in which you'd expect different populations of the same species to follow the same trajectory.
Crampton's results influenced people like Dobzhanky, Mayr and Huxley who helped to re-establish Darwinism as the principal theory of evolution in the Modern Evolutionary Synthesis. But Crampton also predicted arguably the most important development in evolutionary theory since the modern synthesis. In the middle of the 20th century evolutionary genetics was defined by a single debate. The "classical" school held that populations in the wild would have almost no genetic variation, because for every gene there would be one 'best' version and every member of the population would have two copies of that gene. Arguing against the classical school, the "balance" school argued that, quite often, there would be no single best gene and organisms would do better having two different versions of the same gene**. The ballancers thought natural selection would keep lots of different versions of maybe 10% of a species' genes. Both schools assumed natural selection was such a pervasive force that selection would dictate the way populations were made up, they just disagreed on what would result from it. Here's the funny thing, they were both spectacularly wrong. When scientists started being able to measure he genetic diversity of populations in the 1960s it became clear almost every single gene had multiple different versions. Now, in the post-genomic age there is a database with 30 million examples of one sort of genetic variant amongst humans.
Faced with the overwhelming variation he recorded in partulid shells, Crampton had argued natural selection didn't have a damn thing to do with it. Snails isolated from each other by a mountain weren't adapting to their local habitat, they just varied with respect to traits that had no influence on their survival. The fact two populations were isolated meant each would follow its own path and two populations could drift apart from each other. Faced with the overwhelming genetic variation coming from studies in the 1960s Motoo Kimura proposed the neurtral theory of molecualr evolution. Kimura's explanation was the same as Crampton's, almost all of the variation we see at genetic level has no bearing on the success or failure or organisms so the frequency of different variants drifts around at random. The neutral theory is at the heart of a lot of modern evolutionary genetics, and Crampton had understood the underlying principle 50 years before we knew we needed it!
At the end of his monograph on the partulids of Moorea, Crampton said he'd got as far as his measurements could take him, and it was time for someone to study their genetics. In took a bit longer than Crampton might have hoped, but in the 1960s two leading geneticists took up the study of his snails. James Murray from Virginia and Bryan Clarke from Nottingham spent almost 20 years working in what they called, in more than one paper, the perfect "museum and laboratory" in which to study the origin of species. Their work helped scientists understand, among other things, how ecology can contribute the formation of new species and what happens to species when they hybridise with others from time to time. Then, in 1984, Murray and Clarke had to write the most heart-breaking scientific paper I've ever read. It's written in the careful prose scientists use to talk to each other, but the message it delivered was devestating:
In an attempt to control the numbers of the giant African snail, Achatina fulica, which is an agricultural pest, a carnivorous snail, Euglandina rosea; has been introduced into Moorea. It is spreading across the island at the rate of about 1.2 km per year, eliminating the endemic Partula. One species is aiready extinct in the wild ; and extrapolating the rate of spread of Euglandina , it is expected that all the remaining taxa (possibly excepting P. exigua) will be eliminated by 1986-1987.


The bad guys: Euglandina on the left, Achatina on the right.
Euglandina rosea is better known as the Rosy Wolf Snail. It senses the mucous trails of other snails, tracks them down and eats them. It's not clear if the wolf snail had any effect on the pest species it was introduced to control, but it had huge impact on the partulids. By the time Murray and Clarke wrote their paper, E. rosea had already done for one species and it was too well established to control. All they could do was watch as human stupidity and molluscan hunger slowly (1.2 km per year) destroyed the species they'd been studying for 20 years and Crampton had dedicated 50 years of his life to. The same slow torture played itself out in Tahiti and then the rest of the Society Islands. Where there were 58 named species, there are now 5 alive in the wild. Crampton's hundreds of pages of tables should have been the starting point from which the evolution of the partulids could have been tracked. Murray and Clarke's natural laboratory should still be open and be taking advantage of a new generation of technologies that might be able to reveal the genetic and genomic changes that occur when a new species arises. Extinction is a natural part of life, and the fate of all species eventually, but when it's driven by human short-sightedness and robs us of not just a wonderful product of nature but a window through which we might have understood nature's working it's very hard to write about.
I should end by saying there is just a tiny scrap of good news in this story. The partulids are no longer an iconic species in the study of evolution, but they have become the pandas of invertebrate conservation. Murray and Clarke were able to get 15 of the species of the islands and into zoos and labs across the Northern Hemisphere. Breeding programs have been succesful, and new lab-based studies come out form time to time. The relict populations back in the Societies don't have nearly the range they used to, but it appears they've held on to most of their original genetic variation. Perhaps, one day, Eulglandina can be taken care of and some of the partulids can have their islands back.
*Darwin had to be interested in dispersal. Before evolution was widely accepted naturalists thought creatures were created for their habitat (the modern creationist notion of a post-flood diaspora explaining the distribution of animals is almost entirely an invention of Seventh Day Adventists, no, really, it is), Darwin's theory did away with special creation but still needed to explain how life came to live everywhere
** A concept similar to "hybrid vigour", in which crosses between relatively unrelated strains/cultivar bring together different genes and do well as a result. You've seen evidence of this phenomenon any time you've eaten yellow and white "honey and pearl" corn. That corn is a hybrid between a white and yellow cultivar and if you count up the kernals you should get close to the 3:1 ratio Mendel preficts for a dihybrid cross.
Some further reading:
Stephen Jay Gould, who was a snail man himself, wrote and essay on Crampton and the Society Island partulids in which made a humanistic argument for the importance conservation. I resisted the urge to re-read it in researching this piece so anything I stole from him I stole sub-consciously!
- Gould, S.J., 1994. Unenchanted evening in Eight Little Piggies: Reflections in Natural History
Crampton's monograph on the Mooeran partulids (from which the figures above are taken) is available online
- Crampton, H.E., 1932. Studies on the variation, distribution, and evolution of the genusPartula. The species inhabiting Moorea. Carnegie Institution of Washington, 410, 1335.
Finally, the paper in which Clarke and Murray told the world about the demise of their snails:
- Clarke, B., Murray, J. & Johnson, M.S., 1984. The extinction of endemic species by a program of biological control. Pacific Science, 38(2), 97-104.
Labels: environment and ecology, molluscs, Pacific, partulids, sci-blogs, snails, sunday spinelessness

All content not otherwise marked is licensed under a Creative Commons Attribution-ShareAlike 3.0 Unported License.

