Sunday, February 28, 2010
Sunday Spinelessness - Arachnophobia
If you read The Atavism regularily you may have gathered I'm quite fond of spiders (1, 2). So, by and large I'm with the Bug Chicks when if comes to arachnophobia. -No spiders will go out of there way to attack people and there's only a handful of species with toxin that packs any punch to humans so, rationally, there is really nothing to worry about. In keeping with that policy, when this handsome Cambridgea male turned up in our bathroom I saw it as a photo opportunity.

Then he ran cross my camera, up my arm and lodged himself at home on my neck. All my affection for spiders and rationality with regards to the risks lasted about a tenth of a second. I didn't actually scream but it's fair to say my heart rate was somewhat elevated and my movements were restricted. But why? What is is about spiders that freaks us out so much? I did a bit of digging through the academic databases but psychology really isn't my science and I couldn't get a clear idea. Researchers in Germany* have shown (at least among German undergraduate students) spiders do have a special place in our fears - people are more likely to respond badly to photos of spiders than other arthropods, even hymenopterans which can prose a more serious threat to people than spiders. But the question of why remains. Evolutionary psychologists have suggested arachnophobia might be an adaptation but it's hard to imagine that selective pressure applied by occasional spider bites would be sufficient to drive a specific fear of them. Perhaps lots of spiny legs, beady eyes and fearsome fangs just set off enough triggers in the brain to elicit a unique response (did I mention psychology's not my science...).
I remember reading a more plausible (and even testable) idea about the origin of arachnophobia in one of Steven Pinker's books that didn't pop up in my search of the literature. I don't have the book in front of me (and I can't even remember which one it was, although it sounds like a Blank Slate kind of an idea) so you might want to take this with even more grains of salt than you usually would with adaptationist ideas. From memory the argument went that we may be born with a general fear of all spiders and snakes but, until recently, would have learnt the few that are actually dangerous in the area we grew up in from our families. With the good guys and the bad guys separated we could stop wasting our energies on being worried by the overwhelming majority of species which are benign. An inbuilt arachnophobia with inbuilt malleability to surroundings. In the West we aren't exposed to many spiders of any sort in our youth so we retain the childish arachnophobia all our lives. It's a nice story whether it's true or not doesn't seem to have been tested.
Oh, and if my photo of a Cambridgea didn't put you off Te Ara has a much scarier one.
*Spiders are special: fear and disgust evoked by pictures of arthropods. Antje B.M. Gerdes, Gabriele Uhl, Georg W. Alpers. Evolution and Human Behavior, DOI: 10.1016/j.evolhumbehav.2008.08.005)
Labels: arachnophilia, arachnophobia, Cambridgea, environment and ecology, lynx spiders, photos, sci-blogs, sunday spinelessness
Friday, February 26, 2010
Nucleotide diversity - what two new African genomes mean
As I've said before there really is no such thing as the human genome. There are millions of differences between individual genomes and we are each born with about 150 new muations. In an age in which we can sequence assemble and analyse entire genomes in two years understanding the breadth of human genetic diversity is at last an achievable goal and if you want to understand human diversity then you need to look to where we came from. Trace any family tree back far enough and you will end up in Africa and, in fact, most of human history was played out entirely in that continent. Modern humans arose in Africa about 250 000 years ago and only spread out to Europe and the rest of the world in the last 60 000 years, displacing Homo erectus in the process. The migrants that founded the modern European, Asian and American populations would have carried with them only fraction of humanity's genetic diversity when they left Africa but untill recently genomics has focused on those populations. Until last week the two African genome sequences available to researchers were both from Yoruban volunteers to the hapmap project. Although those sequences are very useful they represent only one tip in the deeply branching tree of humanity

To broaden our understanding of African genomes Schuster et al looked to the South of the continent and at two people in particular. !Gubi is a Khoisan (or bushman), a member of a one of the earliest diverging groups within the humanity while Desmond Tutu hails for various Bantu peoples. The results taken from theses genomes along with lower density sequencing and genotyping of other Bantu and Khoisan volunteers reinforces just how much genetic diversity exists within Afirca. By using a method called principle component analysis to reduce a the correlations among millions of single base pair differences (single nucleotide polymorphisms or SNPs) to a smaller set of uncorrelated vectors you can see patterns in the genetic diversity of groups. Applying this method to West African (Bantu and Yoruba), Khoisan and European populations reveals the comparative genetic homogeneity within Europeans and that the difference between the two African groups is comparable to that between either of them an Europeans.
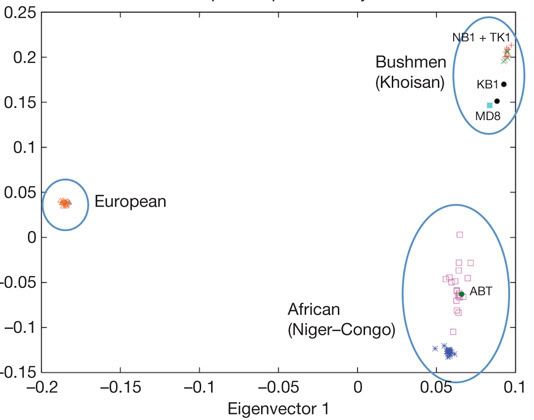
The paper is available to under a creative commons license here and if you feel suitably qualified you can play with their data which has been released on the Galaxy framework.
Labels: evolution, genetics, genomics, Human evolution, human genome, research blogging, sci-blogs, science
You have to be in to win...
Labels: blog blogging, might interest someone, research blogging, sci-blogs
Saturday, February 20, 2010
Sunday Spinelessness - Animals that don't move
There aren't many universal laws in biology. Snails proved Dollo wrong, retorviruses did for Crick's Central Dogma of Molecular Biology and every lesson on Mendel's Laws of Inheritance includes a section of the exceptions to those rules. Biology's disregard of human laws notwithstanding, you might think, at least as far as macro-organisms are concerned, you could safely generalise that animals move and plants stay still. But once you consider the ocean even that generalisation can't be supported; corals, bryozoans, sea squirts, anemones and sponges are all animals that spend their adult life in one spot.
While I was in Masterton for christmas my girlfriend went to Vanuatu (no, you're right, that doesn't quite seem fair...) with a waterproof camera so I've stolen a few of her photos of coral to illustrate todays sunday spinelessness.


It's easy to see why early naturalist thought corals were plants but what you are looking at in those photos is not a single organism, rather it's a colony of tiny genetically identical animals. Corals are members of the phylum Cnidaria (the 'c' is silent) which includes corals, anemones and a diverse bunch of animals we call jellyfish. Such a diverse collection of animals are united under the name Cnidaria because they all employ the impressive nematocyst, a barbed harpoon like cell, to catch and deliver toxin to their prey. Cnidarians have two distinct life stages - a swimming "medusa" (adult jellyfish being the classic example) and a sessile polyp (like the sea anemones familiar to rock pool fossickers the world over). Individual coral colonies (termed "heads") are made entirely of polyps which reproduce asexually depositing a calcium carbonate base as they grow - the exact pattern in which polyps bud from their parents determines the shape a coral head takes.
Many tropical corals supplement their diets by forging a symbiotic relationship with swimming algae (arguably plants that move...) called zooxanthella, the algae get carbon dioxide from the coral's respiration while the coral gets energy from the algae. This relationship is of huge importance in the tropics because it allows corals to grow in those region's warm, nutrient poor waters. Without coral reefs, made from thousands of years of calcification from corals, tropical waters would be nowhere near as biodiverse as they are now and people whose love of animals only goes as far as that peculiar phylum Chordata should care about that:


Labels: anthozoa, cindaria, coral, environment and ecology, might interest someone, sci-blogs, sunday spinelessness
Sunday, February 14, 2010
Sunday Spinelessness - robber fly

I've said before that the bugs in my parents' back yard seemed especially hungry over the Christmas break. Above you see more evidence to this fact. In the lower half of the photo is a robber fly and above is its meal, a blowfly of some sort. I've never seen live robber flies in Dunedin (though there is a dead one here) but the species captured above seems to be reasonably common in the Wairarapa. The first time I spotted one of them I struggled as to where to place it among the insects. The robber flies have long slender bodies and large rounded eyes which misled me into thinking that first sighting might be a small dragonfly. The real taxonomic position of the robber flies is spelt out in bright yellow in the next photo.

That bright yellow structur under the main wing is a haltere. Most insects have two sets of wings and we can tell quite a lot about where a given species fits in the insect scheme based on how it uses those two. Dragonflies and damselflies use both for flying, in beetles the forewings are "sclerotised" into a rigid case that protects the flight wings and the "true flies" ( order Diptera) have turned thier hind-wings into halteres - greatly reduced wings which act as gyroscopes stabilising the flies' flight and allowing them to perform aerobatic tricks. The robber flies from the extremely widespread and speciose family dipteran family Asilidae which includes a staggering 7 000 described species, meaning there are rather more robber fly species in the world than there are mammalian ones.
The features that led me to mistake that first robber fly that I saw for a dragonfly are likely the result of convergent evolution - dragonflies and robber flies are both predators that specialize in taking other insects on the wing. The robber flies differ from odonates in having piercing mouth-parts which they use to inject first a neurotoxin then digestive enzymes into their prey. The blowfly in the photos in this post is paralysed and its tissues are in the process of being liquified and sucked through the robber flies mouth parts. But ever before the neurotoxin entered the blowfly's body it was done for, the robber fly's strong "raptorial" legs are covered in sharp spikes and end in claws that offer little hope for escape once a catch is made.

Labels: Asilidae, diptera, environment and ecology, might interest someone, photos, sci-blogs, sunday spinelessness
Friday, February 12, 2010
Charles Darwin and the Origin of Spouses


Happy Darwin Day everyone! Today would have been Charles Darwin's 201st birthday so around the world geeks are celebrating, churches are standing up to creationism and at least a few biologists are trying to eat their way through the tree of life. With Darwin Day falling so close to Valentines Day I thought it might be fun to forget about Darwin's science just for a few minutes and look at his attitude to love and marriage.
No one has ever accused Darwin about making a rush to judgement about any topic. Just as he spent years poring over the minutest detail of barnacle anatomy before he published The Origin he gave the topic of marriage careful consideration before singing on. In fact, preserved in his notebooks we have a record of the deliberations he undertook. Sometime in 1838 Darwin turned to a new page in his notes and drew a line down the middle, he added the headings "Marry" and "Not Marry" to either side of the line an proceeded to list the pros and cons of either decision. You can see the notebook here but below (presented without comment) is a transcript :
Marry
- Children — (if it Please God)
- Constant companion, (& friend in old age) who will feel interested in one
- Object to be beloved & played with —better than a dog anyhow.
- Home, & someone to take care of house
- Charms of music & female chit-chat.
- These things good for one's health.
- Forced to visit & receive relations but terrible loss of time.
Not Marry
- No children, (no second life), no one to care for one in old age.
- What is the use of working 'in' without sympathy from near & dear friends—who are near & dear friends to the old, except relatives
- Freedom to go where one liked — choice of Society & little of it.
- Conversation of clever men at clubs
- Not forced to visit relatives, & to bend in every trifle.
- To have the expense & anxiety of children
- Perhaps quarelling
- Loss of time.
- Cannot read in the Evenings
- Fatness & idleness
- Anxiety & responsibility
- Less money for books &c
- If many children forced to gain one's bread. (But then it is very bad for ones health to work too much)
- Perhaps my wife wont like London; then the sentence is banishment & degradation into indolent, idle fool
- My God, it is intolerable to think of spending ones whole life, like a neuter bee, working, working, & nothing after all.
- No, no won't do. — Imagine living all one's day solitarily in smoky dirty London House.
- Only picture to yourself a nice soft wife on a sofa with good fire, & books & music perhaps — Compare this vision with the dingy reality of Grt. Marlbro' St.
When I am with you I think all melancholy thoughts keep out of my head but since you are gone some sad ones have forced themselves in, of fear that our opinions on the most important subject should differ widely. My reason tells me that honest & conscientious doubts cannot be a sin, but I feel it would be a painful void between us. I thank you from my heart for your openness with me & I should dread the feeling that you were concealing your opinions from the fear of giving me pain.
Labels: Darwin, Darwin Day, evolution, history of science, might interest someone, sci-blogs
Sunday, February 7, 2010
Sunday Spinelessness - now with 150% more links
None of my photos today, instead I thought we should have a look at what some of web's other inveterate invertebrate bloggers have been up to.

Giant Microbe's plush Tardigrade
- Ted MacRae at Beetles in the Bush hosted the latest edition of the Circus of the Spineless which includes posts on noisy ants, maggot therapy, the place of amphipods in pop culture and no fewer than two samurai related stories.
- Weird Bug Lady (check out her Etsy shop, it's amazing...) now sells a stuffed toy in the shape of a flatworm. Yes, plush planarians!
- Sticking with the plush theme (as phrase I never thought I'd write...) the people behind the wonderful Giant Microbes have stretched their reach into Metazoa, the latest example a plush water bear (Tardigrade)! I note they are not yet available in the Australian/ New Zealand version of their store, but how can animal this cool fail to become a top seller?
- There's yet more invertebrate craft at I Love Insects where Erika finds oragami insects
- Finally, Mike at Arthropoda has more evidence of the coming insect revolution - bees have learned how to recognise our faces. I for one refuse to make the obvious joke.

Weird Bug Lady's plush planarians
Labels: might interest someone, sci-blogs, sunday spinelessness
Thursday, February 4, 2010
Did the Moa's ancestor fly to New Zealand?


The group of birds to which the moa belonged, the ratites, have long fascinated evolutionary biologists. All the ratites are flightless (though, as we'll see they are related to the quail-like tinamous which can fly passably well) and all the major landmasses that had their start in Gondwana had at least one ratite species before humans arrived on the scene. Africa has ostriches, South America the rhea, Australians have emus and cassowary, Madagascar had the Elepahant bird and New Zealand lost the moa but retains the kiwi. The far flung distribution of the ratites and their apparent lack of ability to disperse between continents has led to them being put forward as a classic example of an idea called vicariance biogeography in which the evolutionary history of a group is driven by the geological history of the land on which they live.
For vicariance biogeographers the evolution of the ratites was driven by the movement of the continents. The ancestor of all modern ratites was a flightless bird living in Gondwana and as each new continent split and rifted away from the super continent it took with it a population of ratites which adapted to the ecological changes brought on by their continent's journey: cassowaries in the Wet Tropics of Australia, ostriches on the African Savannah, rhea on the Pampas. It's certainly a nice story, but science has a way of ruining nice stories. The role of vicariance of evolution in the ratites was put to the test once we became able to use molecular evidence to reconstruct the relationships between species. If the geologically driven sketch of ratite evolution I presented above is right then the pattern of branching we find among ratites from different continents should match the order in which we know the continents broke up, something like this:
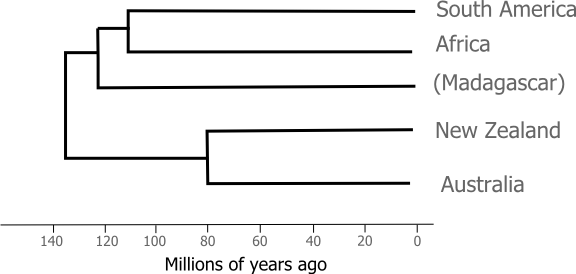
In 2001 Alan Cooper and colleagues sequenced the entire mitochondrial genome (some 12 000 bases of DNA) from representatives of each of the extant ratites and, remarkably, two species of moa. The long, careful process of retrieving DNA sequences from sub-fossil bones deserves a post of its own but for the sake of this article we only need to know what Cooper et al found when they used that DNA to recover the the relationships between ratite species.

The species in bold text above don't fit the pattern that we'd expect from geology alone. If ratite relationships simply reflected the Gondwanan breakup we'd expect to see ostriches grouping with rheas (and apart from the other ratites). New Zealand's two ratite orders are even more surprising, the kiwi lineage is more closely related to the Australian ratites than it is to the moa species. When combined with a molecular clock analysis Cooper et al. concluded that modern kiwis are the descendants of ancient immigrants hailing from either Australia or islands in the Lord Howe Rise (which have since submerged). In order to explain that trans-tasman dispersal the authors reached for the last resort of the desperate biogeographer and invoked a land bridge for which there is little geological evidence. In fact, as we'll see it now seems more likely that the ancestors of the kiwi and the moa flew to New Zealand.
Even with the mitochondrial phylogeny of the group published there was considerable room for uncertainty in how the ratites related to each other. The underlying shape of ratite tree makes it particularly difficult to accurately recover with phylogenetic methods. When we use DNA sequences to estimate a phylogenetic tree we need to find species that share mutations that have accrued during the evolution of the group we're looking at. The branches that relate the different ratite species are relatively short, so there was little time for mutations that set related groups apart from more distantly related ones to accrue. Even worse, the branches that reach to the modern species (the tips of the tree) are very long meaning there has been a lot of time to any mutations that did accrue in those critical short branches to be overwritten*. There are three approaches to dealing with this problem - sequence more genes (since each unlinked gene acts as a separate witness to the evolution of the group), sequence more samples (especially if doing so breaks up a long branch) or use a better model for the way mutations accrue in the genes you are studying. People have tried all three methods to get a better look at ratite evolution. Last year a group centred around the Field Museum in Chicago published a mutli-gene phylogeny of all birds that contained a big surprise for ratite evolution- the most recent common ancestor of all ratites flew.

As long as the ratites grouped together in a phylogeny it was reasonable to assume that they all inherited their flightlessness from the common ancestor of the group. The Field Museum study found that, in fact, the flying tinomous fit right in the middle of the flightless ratites. So, either the most recent common ancestor of the ratites and the tinamous flew and ratite lineages have subsequently lost that ability at least three times or that ancestor was grounded and the tinamous have rediscovered flight. In vertebrates the evolution of true flight has happened three times (in bats, pterosaurs and birds) while there are hundreds of examples of birds that have given up on flying. Moreover, a group of flying birds that are prone to flightlessness is hardly anything new - at least 30 species of rail (including our own weka and takahe) have taken to life on the ground. Given the ways the odds are stacked towards losing flight it seems probable the common ancestor that relates tinamous and ratites flew. The Field Museum study didn't include any moa species and didn't attempt any molecular dating so it's hard to see just how the ancestors of the kiwi and the moa made it to New Zealand
A new study (I knew I'd get to it eventually) published in Systematic Biology throws some light on the New Zealand ratite story. Matt Philips and a team of researchers from the Alan Wilson Centre at Massey University took another look at the mitochondrial dataset used in Cooper et al's 2001 study by adding more kiwi species and using models of DNA evolution that avoid some of the pitfalls of the ratite phylogeny's difficult shape. The new ratite tree and a molecular clock analysis based on that tree confirm the idea of multiple loses of flight in the ratites and add a new finding - the closest living relatives of our giant moa are the quail-like tinamous:
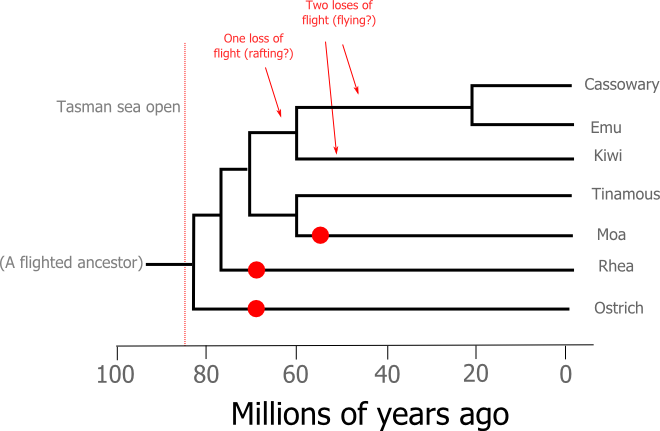
So what does the new understanding of ratite relationships mean for our ideas about the origins of New Zealand's ratites? The molecular clock doesn't quite tick with the regularity of a stopwatch, so there is a good deal more uncertainty in the timing of the events presented above than the precisely defined nodes suggest. Still, even with the uncertainty of molecular dating taken on board we can safely say that both the New Zealand ratite lineages departed from their closest relatives after the Tasman Sea opened up.
The revelation that the tinamous are the moa's closest living relatives suggests that the moa had ancestors that could fly. So, it seems the first proto-moa to arrive in New Zealand flew, or more likely was blown, here from Antarctica. Antarctica? It still seems amazing to me but 30 million years ago Antartica was still attached to South America and, without the circumpolar current to isolate from the world, was a relatively verdant continent. We know from fossils that Antarctica supported southern beech forests (still found in Chile, New Zealand and Australia) and marsupial mammals (strangely absent in New Zealand but still present in Australia and South America) so it's no great stretch to propose the representatives of another Gondwanan group lived there. Antarctica certainly seems like a more likely jumping off point for dispersing proto-moa than South America, but either way it certainly seems they made it here under their own power.
The mode of dispersal for the kiwi's ancestor is a little less clear. As we've seen we can be quite sure that they arrived in New Zealand after the Tasman Sea opened up and there is really no good evidence that there was ever a land-bridge across that sea. We can probably rule out walking. If we disregard the problem of dispersal for a second the simplest way to explain the distribution of flightlessness on the Phillips et al phylogeny is with a single loss of flight in an ancestor shared by kiwis and the Australian ratites. Under that scenario the kiwi would, presumably, have had to raft to New Zealand. Alternatively, given that we've seen the ratites seem to have an inbuilt propensity to becoming flightless we might imagine that the common ancestor shared by the kiwi and the Australian ratites could fly and each lineage has since lost that ability. In this case the kiwi could simply have flown from Australia to New Zealand (a journey that storms frequently inflict on Australian birds today). Without sufficiently old ratite fossils from either country it's hard to choose one scenario over the other.Long range dispersal by rafting is probably an important force in biogeography but if I was forced to make a bet I'd put my money on ancient flying kiwis.
The radical rethink of ratite evolution that a decade of molecular phylogenetics has forced on us raises a lot of interesting questions. What it is it about the ratite body plan, development or behaviour that makes them so prone to flightlessness? Is that repeated loss of flight, and consequent lack of pressure to keep their weight down, enough to explain the trend towards gigantism? The authors of the most recent paper suggest both trends might be explained by ratites on each continent filling the ecological niches left by the extinction of the dinosaurs. The dates on their tree are certainly consistent with the idea that each ratite lineage independently took to the ground 65 million years ago but without more fossils and more precise dates for each split it's very hard to test the idea further. I'm sure the story of ratite evolution has more surprises for us to uncover.
Links to the primary literature are provided below but you should also check out Simon Collins excellent piece in the Herald and Mike Dickison, who got his PhD studying giant flightless birds and wrote about the idea that ratites flew to New Zealand way back in 2007.
Cooper A, Lalueza-Fox C, Anderson S, Rambaut A, Austin J, & Ward R (2001). Complete mitochondrial genome sequences of two extinct moas clarify ratite evolution. Nature, 409 (6821), 704-7 PMID: 11217857 Harshman, J., Braun, E., Braun, M., Huddleston, C., Bowie, R., Chojnowski, J., Hackett, S., Han, K., Kimball, R., Marks, B., Miglia, K., Moore, W., Reddy, S., Sheldon, F., Steadman, D., Steppan, S., Witt, C., & Yuri, T. (2008). Phylogenomic evidence for multiple losses of flight in ratite birds Proceedings of the National Academy of Sciences, 105 (36), 13462-13467 DOI: 10.1073/pnas.0803242105 Phillips, M., Gibb, G., Crimp, E., & Penny, D. (2009). Tinamous and Moa Flock Together: Mitochondrial Genome Sequence Analysis Reveals Independent Losses of Flight among Ratites Systematic Biology, 59 (1), 90-107 DOI: 10.1093/sysbio/syp079Labels: biogeography, evolution, might interest someone, moa, new zealand, phylogenetics, ratites, sci-blogs, science

All content not otherwise marked is licensed under a Creative Commons Attribution-ShareAlike 3.0 Unported License.